
More than 700,000 Americans are afflicted with Type 1 diabetes. Type 1 diabetes (scientific name: insulin-dependent diabetes mellitus or IDDM; colloquial name: juvenile diabetes) typically begins before age 40. The peak age of diagnosis in the U.S. is approximately 14.
Researchers from the Children's Hospital and the Howard Hughes Medical Institute, and the Dana-Farber Cancer Institute, using an Advanced Photon Source (APS) beamline managed by the BiosCARS Collaborative Access Team, have discovered important clues that promise to further research into the prevention of IDDM.
IDDM is a chronic condition in which the pancreas makes little or no insulin because the beta cells have been destroyed due to an autoimmune disorder. The pancreatic islet cells that produce insulin are mistakenly seen by the body as an "enemy." The body then creates antibodies (or antigens) to fight the "foreign" tissue. These antigens destroy the islet cells' insulin-producing capacity and the resulting lack of insulin brings on diabetes. IDDM usually presents abruptly, although the damage to the beta cells may begin well before the disease manifests.
Prevention of IDDM requires a thorough understanding of what biological anomolies trigger the autoimmune response against pancreatic islet cells. In order to gain that understanding, the structures and binding mechanisms of protein molecules that produce the antigens must bedetermined. Research has shown that IDDM sufferers carry certain genes coded for cell-surface proteins important in the production of islet cell antigens. These are the class II Major Histocompatibility Complex (MHCII) glycoproteins HLA-DQ8 and HLA-DQ2 in humans and I-Ag7 in non-obese diabetic mice (NOD). In IDDM, autoimmune destruction of pancreatic islets is preceded by the appearance of autoantibodies and autoreactive T cells to a number of islet antigens, including insulin and glutamic acid decarboxylase. The relationship of these autoimmune reactions to the initiation and progression of the disease is still unclear.
X-ray crystallography techniques applied at APS beamline 14-BM-C have determined the three-dimensional structure of DQ8 complexed with an immunodominant peptide (a class of molecules that hydrolyze into amino acids and form the basic building blocks of proteins) from insulin. They have analyzed the size and composition of the peptide binding pockets of DQ8 relative to I-Ag7 and to models constructed of DQ2 and MHCII molecules associated with either IDDM low-risk (DQ7) or dominant protection (DQ6). A marked similarity of the DQ8, DQ2, and I-Ag7 peptide binding pockets suggests that diabetes is caused by the same autoaggressive antigen-presentation event(s) in man and mouse. This discovery provides evidence that the NOD mouse is a good model for the human disease, and can be used as a test subject for research into prevention of IDDM. Correlating IDDM epidemiology and MHCII sequences with the DQ8 structure suggests that other structural features evident from these data contribute to a susceptibility to Type I diabetes.
K. H. Lee,* K. W. Wucherpfennig,Ý & D. C. Wiley*ý
* Laboratory of Molecular Medicine, The Department of Medicine, The Children's Hospital and the ýHoward Hughes Medical Institute, 320 Longwood Avenue, Boston, MA
Ý Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute, 44 Binney Street, Boston, MA
Submitted to Nature Immunology
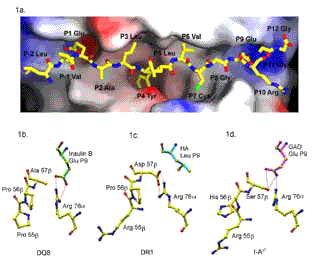
Figure caption:
1a. The surface of DQ8 with InsulinB (9-23) bound (yellow), viewed as seen by a T cell. Peptide sidechains P1-Glu, P4-Tyr, and P9-Glu are buried in deep pockets, while P6-Val and P7-Cys lie on shelf-like surfaces. The P4 pocket of DQ8 is non-polar (white), while the P9 pocket is positively charged (blue). (Red surfaces are electrostatically negative.)
1b. DQ8 P9 pocket with the InsulinB peptide P9-Glu (green) hydrogen bonded to Arg-76[alpha] and with Ala-57[beta] replacing the common Asp-57[beta]. View is from the C-terminal end of the bound peptide (right end of Fig 1a). DQ8 differs from I-Ag7 at [beta]-chain residues 55-57, Pro-Pro-Ala (DQ8) which are Arg-His-Ser in I-Ag7 (Fig. 1d). The [beta]-chain helix residues 55-57 are to the left of Fig 1b-d. Hydrogen bonds are dashed red lines.
1c. DR1 P9 pocket with Asp-57[beta] hydrogen bonded to Arg-76[alpha] and the hemagglutinin peptide P9-Leu (blue) in the electrostatically neutral pocket.
1d. I-Ag7 P9 pocket with Ser-57[beta] and the GAD65(207-220) peptide P9-Glu (purple) hydrogen bonded to Arg-76a and Ser-57b. The b-chain a-helix (56-57) in DQ8 is positioned closer to the Glu-P9 side chain and Arg-76a (Fig. 1b) than in I-Ag7 (Fig 1d). There is not enough space in the structure of DQ8 to replace Ala-57b with the larger sidechain of a Ser as in I-Ag7.